|
| Chapter 10 The Lens NICHOLAS A. DELAMERE Table Of Contents |
The lens is a vital component of the transparent pathway between the outside
world and the retina. The lens and cornea comprise the refractive
mechanism of the eye. However, the lens adds the important element of
variable focus. Because the shape of the lens can be changed, thus altering
its focal length, it is possible for the eye to switch focus from
a distant to a near object. To function properly, the lens must be
transparent. This requires considerable specialization because tissues
do not normally transmit light. Perhaps the most obvious specialization
is the lack of blood supply to the lens. Also, the lens does not have
nerves. It exists in isolation from other tissues, suspended by ligaments
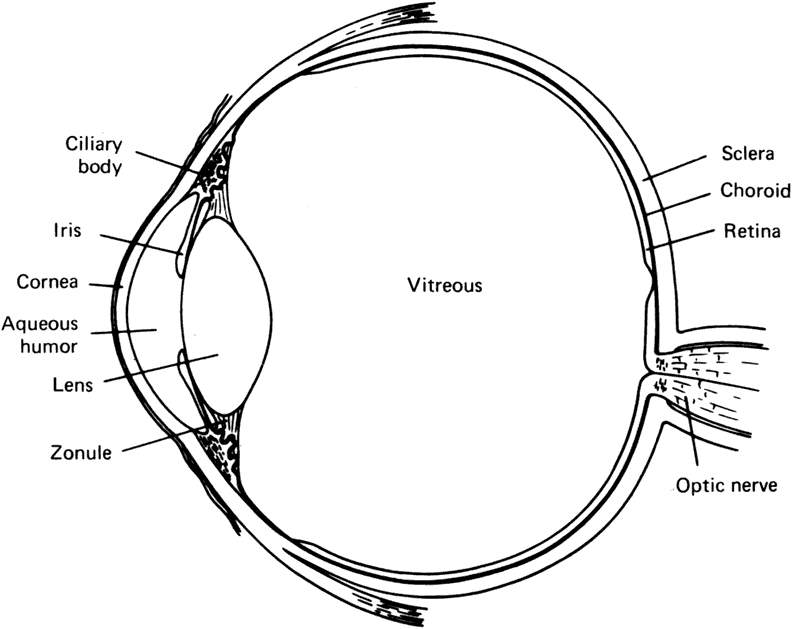
in the fluid-filled interior cavity of the eyeball (Fig. 1). Other, less apparent, specializations include the tight, ordered packing
of lens cells, the loss of organelles in most lens cells, and the
abundant expression of cytoplasmic proteins called crystallins. One consequence
of these specializations is the remarkably long life span of
lens cells and their constituents. Once made, lens cells are retained
for the lifetime of the individual. Because fully differentiated lens
cells are unable to synthesize proteins and lipids, defective or damaged
biomolecules cannot be replaced. Instead, the lens appears to be
highly specialized at preventing protein and lipid damage. It is extraordinarily
efficient at cell preservation, keeping oxidative damage at
a manageable level for decades. This feat is even more remarkable when
one considers that for two thirds of each day potentially damaging light
rays pass through the lens.
|
| FOCUS AND ACCOMMODATION | ||
At rest, the lens in the normal eye focuses images of distant objects on
the retina. It acts in much the same way as a biconvex optical glass
lens (Fig. 2). Thus, the refractive power of the lens is a function of the curvature
of the lens surfaces, as well as the refractive index of the cell mass. The
closely packed, highly ordered array of lens cells is essential
for optical quality, and even small defects in lens fiber cell anatomy
can cause detectable spherical aberration (variability of focal length).1
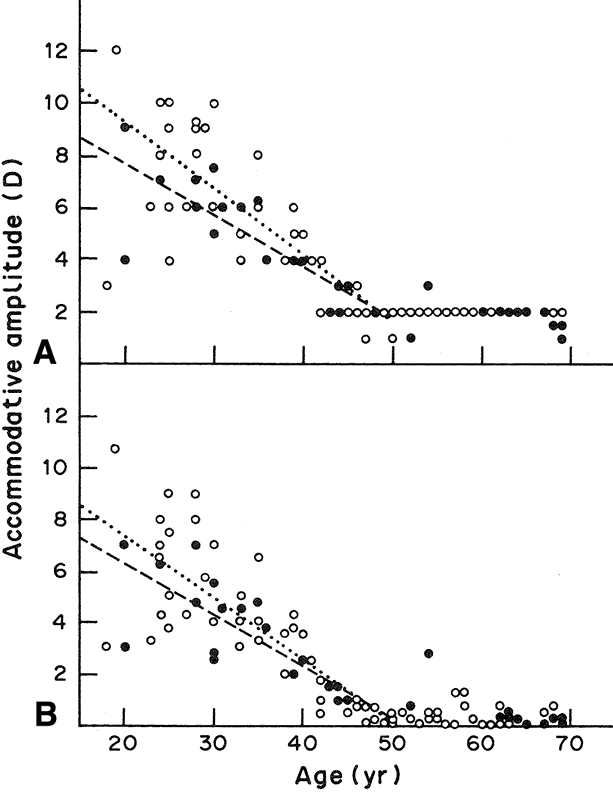
For the eye to focus the image of a near object on the retina, there must be an increase in ocular refractive power. The mechanism that achieves this change is accommodation, and the eye at rest is said to be unaccommodated. The mechanism of accommodation varies among species.2,3 Not all animals accommodate. Some fish retract their lenses to focus on distant objects, whereas in snakes and frogs and many mammals, the position of the lens moves forward for near vision. In certain other fish and horses, the retina is tilted so that the lens-retina distance alters generally in a dorsal-ventral plane. In humans, the ability of the eye to alter focus is facilitated by a change in the shape of the lens; there is no change in the curvature of the cornea or the length of the eyeball. In the human, the accommodative mechanism involves the ciliary muscle, the zonules, and the lens itself. However, the role of the lens is passive in the sense that its shape change takes place as the result of contraction or relaxation of the ciliary muscle. When the ciliary muscle is relaxed, the zonules are under tension, exerting a centrifugal force on the lens equator that causes the lens to take on a flattened form. Contraction of the ciliary muscle makes it shorten and move forward and toward the equator of the lens. As a result, the zonules become less taut; their pull on the lens capsule is reduced; and the lens changes to its relaxed, more spherical, and, thus, accommodated, shape. This results in an increase in the dioptric power of the lens, allowing a near object to come into focus on the retina. To shift the focus back from a near to a distant object, the ciliary muscle relaxes and the elasticity of Bruch's membrane causes it to move posteriorly, widening the ciliary ring so that the zonules become tense once again and flatten the lens. The principle of this currently accepted theory for the mechanism of accommodation was first suggested by Helmholtz4 in 1855, and the important role of the lens capsule was emphasized by Fincham5 in 1937. During accommodation, the principal change in the lens shape is seen at the anterior surface. The anterior surface of the unaccommodated lens has a spherical radius of curvature of about 12 mm. When the eye accommodates, the anterior surface tends to bulge centrally, attaining a radius of curvature of about 3 mm. The more peripheral anterior surface shows relatively little or no increase in curvature. Minimal changes occur in the curvature of the posterior lens surface; Fincham5 detected changes from 5.18 to 5.05 mm in one case and from 5.74 to 4.87 mm in another case for an accommodative change amounting to 8 diopters. The axial thickness of the lens increases during accommodation and the diameter decreases. The increase in lens axial thickness is paralleled by a shallowing of the anterior chamber of the eye. The amount that an eye can alter its refractive power between focus on a far object and a near object is called the amplitude of accommodation. The amplitude of accommodation decreases progressively with age, a condition known as presbyopia6 (Fig. 3). The decline in accommodative amplitude means that the near point becomes more distant from the eye and, thus, small objects must be held farther away to be seen clearly. It is doubtful that there is a single cause of presbyopia. Theories proposed to explain the development of presbyopia include changes in the elastic properties of the capsule, a change in the deformability of the lens cell mass, loss of elasticity in Bruch's membrane, and weakening of the ciliary muscle.7–9 There is little evidence to support the concept of a weakened ciliary muscle, although age-related morphologic changes are known to occur in this tissue.10 In presbyopic eyes, the ciliary muscle is still able to contract vigorously under the influence of pilocarpine.11 Presbyopia does not appear to be simply the result of changes in the rate of lens thickening or anterior chamber shallowing with age.12 Changes in the aging capsule have been studied by Fisher, 13 who concluded that reduction in capsular elasticity in the senile lens is a contributing factor in presbyopia. The principal cause of presbyopia was once considered to be “lenticular sclerosis,” a progressive hardening of the lens with age so that it becomes less deformable. However, in his study of lens mechanics, Fisher14 determined that age-related changes are not entirely consistent with the timetable for the onset of presbyopia.
|
| LENS DEVELOPMENT | |||
The ability of the lens to function so efficiently as an optical instrument
depends to a large extent on its unique anatomy. To understand lens
structure it is helpful to consider its embryonic development. The
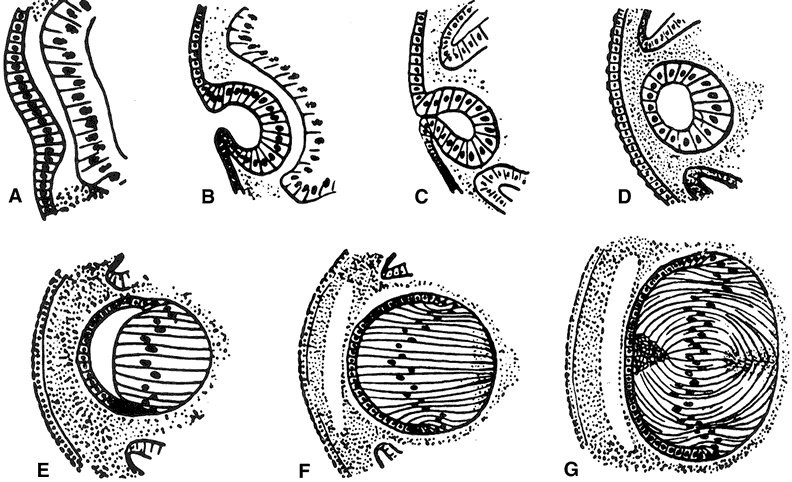
lens plate (or placode) arises from a thickening of the surface ectoderm
of the head fold induced by contact with the optic vesicle, an outgrowth
of the forebrain; this event takes place at the 4-mm stage (4-weeks
gestation) in human embryos (Fig. 4). At about 5-weeks gestation (5-mm stage) the lens plate begins to invaginate
to form the lens pit; at the same time the optic vesicle invaginates
to form the optic cup where the embryonic lens is located. The
lens pit continues to invaginate, forming a structure called the lens
vesicle that separates from the surface ectoderm at approximately the 9-mm
stage (5- to 6- weeks gestation). A thin lens capsule, the basal
lamina, develops around the cell mass at about week 6. Also at around 6-weeks
gestation, the embryonic lens is enveloped by a delicate vascular
system, the tunica vasculosa lentis, which is most extensive at 10- to 12-weeks
gestation and is supplied by the hyaloid artery of the retina. These
blood vessels presumably cater to the nutritional needs of
the developing lens.
The cells of the lens vesicle are positioned so that their apical surfaces face inward and their basal surfaces are in contact with the lens capsule. The cells at the posterior of the lens vesicle elongate and by approximately the 16-mm stage (7-weeks gestation) they have grown to form primary lens fibers that fill the cavity of the lens vesicle. The primary lens fibers eventually lose their nuclei and organelles but what remains of each cell is retained throughout life as the “embryonic nucleus” of the lens. Although the cells at the posterior portion of the embryonic lens grow to become primary lens fibers, the cells at the equator have a different fate. They divide and give rise to new cells. In most cases the newly formed cells differentiate and elongate to form secondary lens fibers. A few of the newly formed cells probably remain as undifferentiated epithelial cells, populating the anterior epithelial monolayer as the lens grows. The equatorial or germinative zone of the anterior lens epithelium continues to divide and produce secondary lens fibers throughout life. It is the only region of the adult lens where the cells remain capable of mitosis. Neither lens fibers nor the central anterior lens epithelium undergo cell division. As the cells in the equatorial zone of the epithelium elongate, the portion of each cell that is anterior to the nucleus grows forward under the undifferentiated epithelium, lying just beneath the anterior capsule; the posterior portion of each growing lens fiber simultaneously elongates backward beneath the posterior capsule. When the ends of similarly elongated cells meet each other close to either the anterior or posterior pole of the lens, both cells stop growing. New fiber cells are formed continually throughout life and older cells are displaced toward the center of the lens. Like tree rings, the youngest cells are at the lens periphery, the oldest at the center. Understandably, the lens center becomes denser. It is referred to as the lens nucleus. The more superficial layers of younger fiber cells are called lens cortex (see diagram, Fig. 5). An interesting feature of the differentiation of lens epithelium into lens fibers is that as the fibers are displaced to the interior of the lens, they reach a stage at which they lose their nuclei and other cytoplasmic organelles.
The zonular fibers, which are suspensory ligaments that support the lens, develop from the neuroepithelium at approximately the 65-mm stage (3-months gestation), when the ciliary body and iris begin to develop from the optic vesicle. At the 110-mm stage (4-months gestation) the zonules are well developed, running from the inner surface of the ciliary body to the lens capsule. Still later, at the 240-mm stage (7-months gestation), the vascular hyaloid system within the vitreous regresses; it is absent by 8.5-months gestation, leaving the lens without a blood supply for the rest of its life. The regulatory factors involved in lens development are complex and have been nicely reviewed by McAvoy and colleagues15 and van Doorenmaalen.16 Lens induction depends on the interaction between presumptive lens ectoderm and other tissues, particularly the retina. Transcription factors Pax-6, as well as Eya, Six, Sox, and Prox play an important role.15 The communication between developing tissues is a two-way street; signals emanating from the lens have a significant influence the development of other parts of the eye17 (Fig. 6). The involvement of specific lens-inducing substances in lens differentiation seems certain.15,18,19 Chamberlain and McAvoy19 have suggested that fibroblast growth factor promotes lens cell differentiation, and Beebe and colleagues18 demonstrate a role for insulin-like growth factors.
|
| GENERAL STRUCTURE OF THE LENS | ||
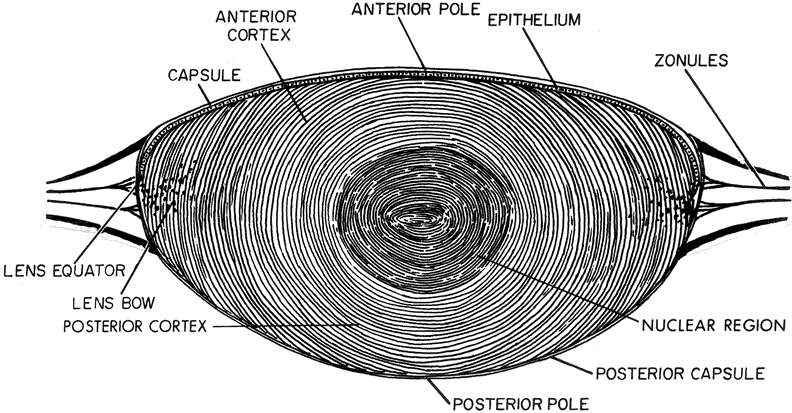
| The lens of the fully developed eye is a biconvex, transparent structure
located immediately in front of the vitreous body and behind the iris (see Fig. 1). The lens of the young eye is generally colorless, but a yellowish to
amber color develops with age. The front surface of the lens, facing
the cornea, is bathed with aqueous humor, which flows through the posterior
chamber, through the pupil, and into the anterior chamber. The entire
cellular mass of the lens (the epithelium, cortex, and nucleus) is
contained within the elastic capsule, which has a smooth outer surface (see Fig. 5). The lens is held in place by suspensory ligaments (zonules) running
from the ciliary body and inserting into the superficial lens capsule
around the equator. The lens is also supported by its immediate apposition
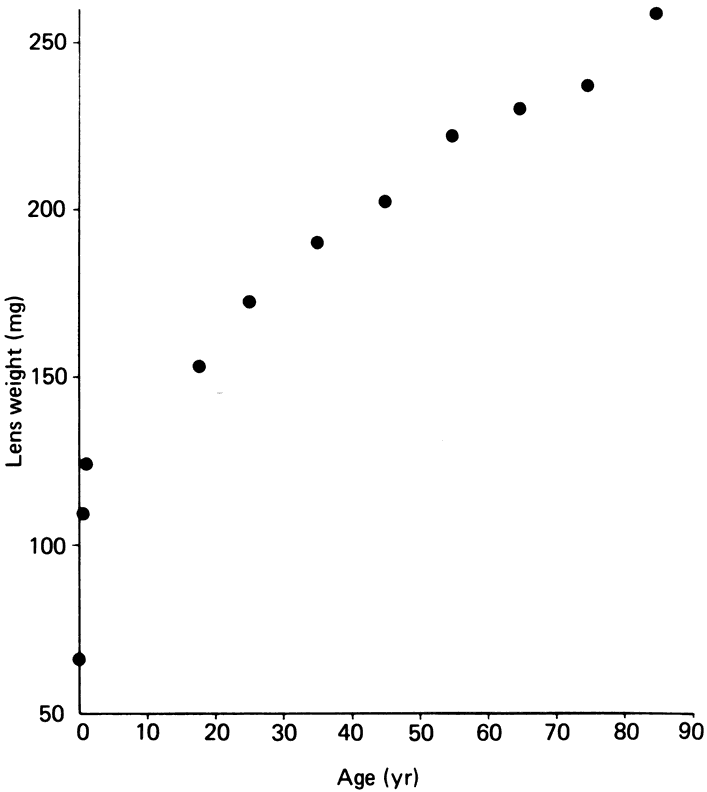
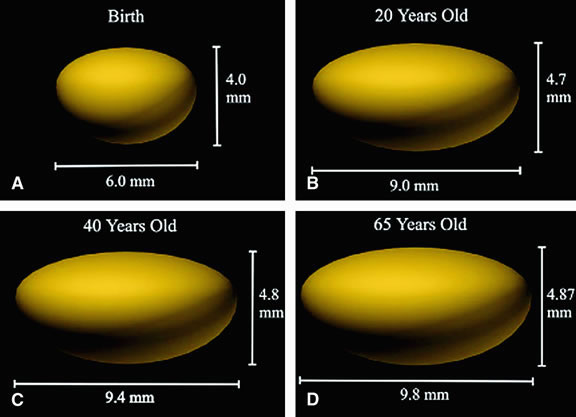
to the vitreous. Cell division and, thus, growth of the lens continue throughout life; as new lens cells are formed, the older cells are displaced toward the interior of the lens. As a result, the lens grows in size and weight as humans age20 (Figs. 7 and 8). It has been estimated that the human lens thickness increases at about 0.02 mm per year.21,22 The anterior-posterior dimension of the lens in the newborn is 3.5 to 4.0 mm, reaching 4.75 to 5.0 mm at 90 years of age. The equatorial diameter in infants is about 6.5 mm, increasing to 9.0 with age. The radius of curvature of the anterior surface of the lens is 8 to 14 mm, and at the posterior surface it is 4.5 to 7.5 mm. There is, however, variation in curvature with age and accommodation.
|
| CAPSULE | |
The cell mass of the lens is completely enclosed by a transparent elastic
acellular capsule. In the adult, the thickness of the lens capsule
is approximately 13 μm at the anterior pole but is considerably thinner (approx. 4 μm) at the posterior pole.23,24 The anterior capsule thickens with increasing age, but the thickness of
the posterior capsule changes very little after birth. The dynamics
of capsular growth have been discussed fully by Rafferty25 and by Parmigiani and McAvoy.26 Viewed by light microscopy, the capsule material appears homogeneous. However, electron
microscopy reveals a fibrillar structure with a more
dense outer layer (the zonular lamella) into which the zonules penetrate 0.5 to 1 μm.27 The inner aspect of the capsule makes contact with the basal surfaces
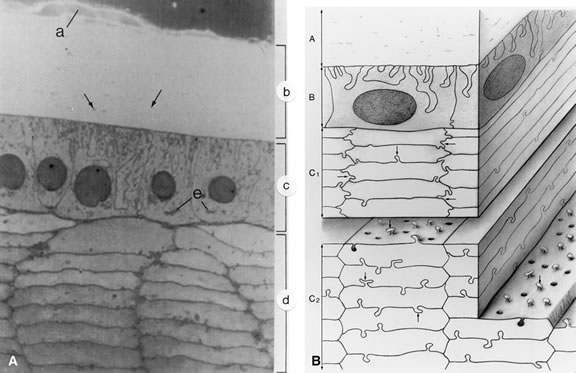
of the epithelial cells anteriorly (Fig. 9) and the elongated lens fiber posteriorly. Electron microscopic studies
reveal a smooth contour on all surfaces except at the equator where
the tension of the zonules produces irregularities.
The lens capsule is analogous to a basement membrane. It is composed mainly of type IV collagen combined with about 10% glycosaminoglycan.28–30 As with other basement membrane material, the lens capsule stains positive with periodic acid-Schiff reagent in histologic section. The capsule is also easily digested by collagenase. Autoradiographic studies by Young and Ocumpaugh31 demonstrated that the capsule is synthesized by the lens epithelium and superficial posterior lens fibers; synthesis of capsular material appears to persist throughout life at the anterior of the lens but not at the posterior surface.26 The zonules, or suspensory ligaments, are anchored in the lens capsule. The zonules hold the lens in place and influence the shape of the lens by the degree of tension they exert. The zonules are structurally and biochemically similar to elastic microfibrils in other tissues.32–34 The capsule and zonules play an important yet passive role in the process of accommodation. Their degree of elasticity is important. As described previously, the shape of the lens changes according to the degree of tension applied to the zonules by the ciliary muscle. |
| EPITHELIUM |
| The lens epithelium exists as a monolayer situated beneath the capsule
at the anterior surface of the lens (see Fig. 9). The epithelial monolayer extends to the equatorial region of the lens
but does not continue across the posterior surface. Thus, the outermost
lens cells that face the aqueous humor are epithelial cells, whereas
the outermost lens cells that face the vitreous humor are fiber cells. With
the exception of cells at the equatorial zone, the epithelial
cells are generally cuboidal, about 5.5 to 8 μm high and 7 to 11 μm
wide.35 The lateral surfaces of these epithelial cells are richly interdigitated. Toward
the equatorial zone the epithelial cells tend to be more cylindrical. A
small population of epithelial cells at the posterior of
the equator, the germinative zone, represents the only group of cells
in the mature lens that remains capable of mitosis. It is here that the
cells divide and differentiate into new lens fiber cells. The plasma membrane domains at the lateral and apical aspect of the epithelium have desmosomes for cell adhesion. The epithelial cell plasma membranes have also been shown to have gap junctions,36,37 which facilitate intercellular communication between the adjacent epithelial cells and less often with the underlying lens fiber cells. The role of gap junctions in the physiology of the lens is discussed later. All lens epithelial cells possess a nucleus together with granular cytoplasm, mitochondria, Golgi apparatus, ribosomes, rough endoplasmic reticulum (ER), and numerous small filaments.35,38 Such intracellular organelles are consistent with the competent metabolic function of the epithelial cells. In addition to cell division, the epithelium is responsible for much of the active solute transport activity in the lens and also for secretion of capsular material. Cytochemical studies of lens epithelium membranes have demonstrated the presence of sodium-potassium adenosine triphosphatase (Na, K-ATPase; the sodium pump enzyme) at the lateral and apical surface membranes.39,40 The epithelium contains a high specific activity of many other enzymes, including acid phosphatase and aldose reductase.41,42 In simple terms, lens epithelial cells have the characteristics of “normal” cells. The same is not true for lens fibers. Although they are differentiated epithelial cells, fibers are highly specialized and have several uncommon features. |
| FIBER CELLS | ||
The bulk of the lens is made up of tightly packed fiber cells (Fig. 10). The most superficial, newly made, lens fibers contain organelles and
a nucleus similar to those of the anterior epithelial cells.35,38,43 However, these structures are not retained. As the cells age and are displaced
farther toward the center of the lens, a stage is reached at
which the cell nucleus fragments and disappears completely, together with
other organelles. The ability to get rid of these intracellular structures
is a highly coordinated process that seems to be programmed into
the fiber cell.44 The fully differentiated, mature lens fiber has no nucleus, no mitochondria, and
no ER. The nonparticulate cytoplasm of the mature fiber cells
is a specialization that contributes to the transparency of the lens.
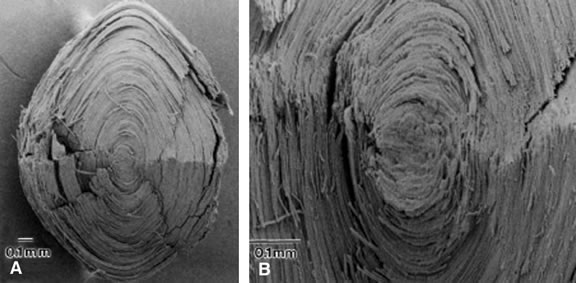
The newly formed elongating lens cells have a hexagonal cross section that they retain throughout life (Fig. 11). Fibers are enormously long but thin. Within the cortical region of the lens, the fiber cells are 8 to 12 mm long, 7 μm wide, and 4.5 μm thick.35 The lens fibers are very densely packed, forming a highly structured, honeycomb-like array with an intercellular distance of only about 20 nm. The intercellular dimension is small compared with the wavelength of visible light, and this, together with the high degree of order, minimizes light scatter and favors transparency. A recently formed lens fiber has its center approximately at the equator, whereas its two ends project forward and backward to the anterior and posterior poles of the lens. If the system were perfectly symmetric, all the fibers would end at the same location at either pole of the lens; this does not happen, perhaps because it would detract from the optical properties of the optical axis of the lens. Instead, the fiber ends meet in complex patterns called lens sutures. The human embryonic lens has Y sutures (λ anteriorly; Y posteriorly), but in the adult lens there is a more complicated four-pointed star arrangement.25
Superficial lens fiber cells show some moderate interdigitations between adjacent cells, but deeper lens fibers have distinct interlocking processes resembling ball-and-socket joints.25,45 Moving farther into the nuclear region, the lens fiber cells retain their hexagonal shape, but the interlocking processes are replaced by ridges. The purpose of these interlocking mechanisms between lens fiber cells may be stabilization of the packing arrangement to prevent slippage of cells against each other when the lens changes shape during the process of accommodation. |
| COMMUNICATION BETWEEN CELLS | |||
Because terminally differentiated lens fiber cells lack a nucleus, mitochondria, and
other organelles, they are incapable of existing in isolation. Their
survival depends on the ability of the lens fiber mass to
function rather like a syncytium, enabling the epithelium and the nucleated
fiber cells at the lens surface to support the nonnucleated fiber
cells underneath. This is made possible by gap junction channels between
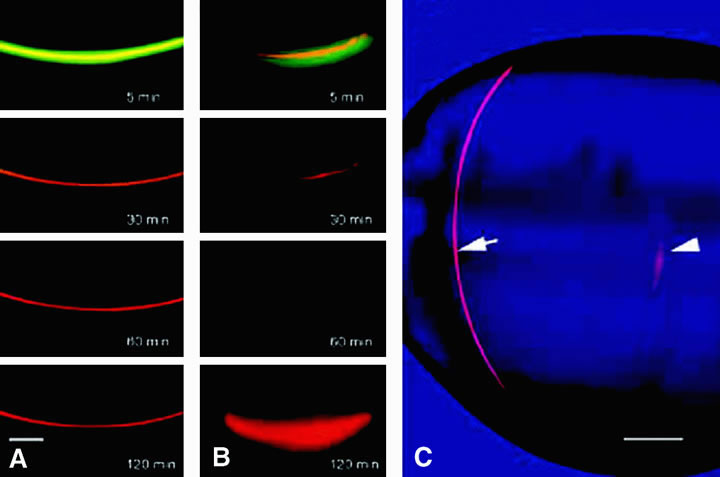
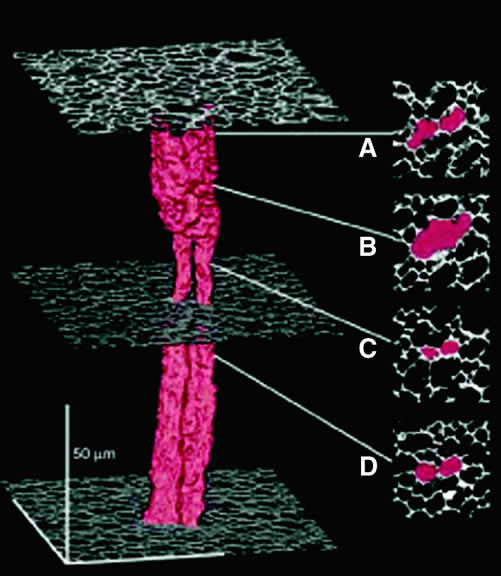
fiber cells (Fig. 12) and possibly also by the occasional fusion of adjacent fiber cells. By
means of these specializations, solutes can pass from cell to cell (Fig. 13). Gap junctions also connect adjacent cells in the anterior monolayer
of epithelium, but junctions between epithelial cells and fibers are sparse.46,47 The gap junctions are formed when hemichannels in adjacent cells dock, creating
a pathway for the passage of ions and low molecular weight solutes. Each
hemichannel is an assembly of connexons that are formed by
specific connexin proteins. More than a dozen connexins are known to
exist, coded by different genes. Connexin-46 (Cx46) and connexin-50 (Cx50) are
found in mammalian lens fibers.48,49 The gap junctions between lens fibers are different from those between
lens epithelial cells in which connexin-43 (Cx43) is predominant.50 The importance of gap junctions is reinforced by the finding that targeted
mutation of either Cx46 or Cx50 in mice results in cataract.49,51 In humans, mutations in Cx46 and Cx50 genes have been associated with
certain congenital cataracts.52,53
The distribution of gap junctions does not appear to be spatially uniform across the fiber cell mass, and the extent of functional cell-cell coupling is high at the equator and low at the anterior and posterior poles.47 Fiber cells located within 400 μm of the lens surface are able to reversibly uncouple in response to stimuli such as a change of pH; the fiber cell connexons can apparently regulate their conductance. More deeply located, older fibers are not capable of uncoupling, and this may stem from posttranslational modification of the connexin proteins or the absence of essential cofactors. There could, however, be other pathways that couple deeply located fibers. It has been found that cell-to-cell fusion occurs in the lens core54 (Fig. 14).
|
| WATER | ||||||||||||||||||||||||
| The lens is unusual in that it has a low water content (65%) and an extraordinarily
high protein content (33%). Almost all of the lens dry weight
is protein. Low water content and high concentration of refractile
protein are essential for the optimal optical function of the lens. The
composition enables the lens to have a refractive index considerably
greater than its fluid environment and yet remain sufficiently hydrated
to be deformable during the process of accommodation. Lipids, inorganic
ions, organic phosphate, nucleic acids, and various metabolites
comprise only about 2% of the total lens solids (Table 1). It should be noted, however, that the composition of the lens varies
with aging. Furthermore, there can be a number of dramatic changes in
lens composition that accompany cataract.
*Solute concentrations are shown as mmol/kg lens H2O.
The low water content of the lens partly reflects the unique protein composition of fiber cell cytoplasm. It also reflects a lack of extracellular space. Although the adult human lens capsule is about 80% water,55 the cortex contains 68.6% water and the nucleus 63.4% water.56 The normal human lens shows no significant alteration in hydration with age.56,57 Not all the water in the lens is freely diffusible. A fraction is involved in protein hydration and is, thus, considered bound. Earlier evidence for different fractions of lens water was obtained using flux studies with radioactively labeled water. Nuclear magnetic resonance (NMR) studies suggest that about half of the lens water is physically associated with lens protein.58 Because the fibers are packed tightly, very little of the lens water is located in the spaces between the cells. The extracellular space of the lens has been measured by determining the distribution of tracer substances (e.g., inulin) that do not penetrate the lens cell membranes. In the rabbit lens, the inulin space has been reported to be about 5%.55,59 Nearly a fourth of the measured extracellular space is located in the capsule.55 Lens hydration can be significantly higher in certain forms of cataract. Precise regulation of lens hydration is critical for the maintenance of lens transparency. Cell swelling, cell shrinkage, or enlargement of the extracellular spaces result in disruption of the highly ordered arrangement of lens fibers, causing light scatter. Fischbarg and his coworkers60 suggest that the lens epithelium is able to transport water inward to the fiber mass. Water may be shifted out of the lens at the equator.47 The expression of specialized water channel proteins, aquaporins, might enable the lens to perform these functions. |
| PROTEIN | ||||||||||||||||||||||||||||||||||||
CRYSTALLINS Lens proteins can be broadly separated on the basis of their solubility in water. The water-soluble component accounts for 80% to 90% of mammalian lens protein and is composed mainly of the lens crystallins. In humans, the major proteins are α-, β-, and γ-crystallin; in reptilian, avian, and invertebrate species there are other crystallins (Table 2). The water-insoluble fraction of lens proteins can be further separated on the basis of its solubility in urea. The urea-soluble fraction contains cytoskeletal proteins and modified crystallins. The urea-insoluble fraction contains mainly membrane protein. The original separation of lens proteins into soluble and insoluble fraction was accomplished in 1894 by Morner.61
*As described by Piatigorsky (1998) some lens crystallins are functional proteins expressed at low levels in other tissues but expressed in high abundance to serve as refractive proteins in lens cells. In different species, the different proteins have been recruited as lens crystallins.
α-Crystallin accounts for as much as 40% of total lens protein. α-Crystallin is not a single protein but a complex or aggregate of 30 to 40 copies of αA-crystallin and αB-crystallin in roughly a 3:1 molar ratio. The αA-and αB-crystallin polypeptides have a molecular mass of approximately 20 kilodaltons (kDa). At the time of synthesis, the αcrystallin complex has a molecular weight of about 7 × 105 daltons (Da). With aging, larger aggregates of α-crystallin are formed with molecular weights as high as 50 × 106 Da; these aggregates of αcrystallin become water insoluble and have a tendency to bind to the plasma membrane of fiber cells.62,63 This might contribute to a decrease in transparency. From a three-dimensional standpoint, about half of the α-crystallin polypeptide chains are in a β-sheet configuration; there is also some β-turn and helical configuration.64,65 The amino acid sequence indicates approximately 55% homology between αA and αB.66 The αA-crystallin gene is an evolutionary sibling of the αB-crystallin gene.67 Although there is much similarity between αA- and αB-crystallin, there are marked differences in their expression. αA-crystallin is almost exclusively found in lens fibers, whereas αB-crystallin is expressed in numerous tissues including heart, skeletal muscle, brain, lungs, and kidney, as well as lens.68 Studies on knockout mice indicate differences in the role of αA- and αB-crystallin. Deletion of αB-crystallin has little apparent effect on lens morphology, whereas αA-crystallin deletion causes cataract.69 Similarity of amino acid sequences suggest that β- and γ-crystallins are products of the same gene family, which is distinct from α-crystallin.65 The β-crystallins are the most abundant of all lens proteins, representing about 54% of the total water-soluble protein. The molecular weight of the β-crystallins ranges from 4 × 104 to 2.5 × 105 Da. The β-crystallins are heterogeneous and can be divided into a number of subgroups. The first four subgroups of aggregates to be identified have molecular weights estimated to be 250,000, 130,000, 60,000, and 37,000 Da66; of these, the two larger species are called β-high and the smaller species are called β-low. Each of the β-crystallins contains two subunits; the major subunit, βBp, has a molecular weight of approximately 24 × 103 Da. The βBp chain is a characteristic β-crystallin polypeptide found in a great number of mammalian species. The three-dimensional structure of β-crystallin is principally a β-sheet configuration.64,70 Although β-crystallins are abundant, the γ-crystallins represent only 1% to 2% of the total lens protein. γ-Crystallins are found in the monomeric form with a molecular weight in the range of 20,000 to 27,000 Da. Like α- and β-crystallin, γ-crystallin has relatively little α-helical structure; its configuration is mainly β-sheet and some β-turn. γ-Crystallin has been implicated in the so-called cold cataract; if the temperature is reduced, the γ-crystallin can undergo a conformational change, resulting in opacification of the lens; with warming, the cold cataract reverses.70,71 It has been known for a long time that crystallins comprise the bulk of lens proteins and that their expression gives the lens unique optical properties. However, the original notion that these proteins are lens specific turned out to be incorrect. Analysis of crystallin amino acid sequences revealed that many of the proteins are either closely related or, in some cases, identical to enzymes and stress (or heat-shock) proteins found in nontransparent tissues67 (see Table 2).Consistent with its identity as a stress protein, α-crystallin is able to suppress the heat-induced nonspecific aggregation of a range of soluble proteins including insulin, α-lactalbumin, and alcohol dehydrogenase. In this respect, α-crystallin is a molecular chaperone. In a variety of tissues as well as in the lens, α-crystallin likely plays a role in ensuring the correct folding of proteins. Evolution has led to gene sharing,67 which is an apparent recruitment of functional gene products that are normally produced in modest amounts elsewhere but are expressed in high abundance in the lens as refractive proteins. MEMBRANE PROTEINS The protein associated with lens cell membranes represents 20% to 30% of the water-insoluble fraction of the lens proteins72; the membrane proteins are insoluble in urea. Membrane proteins are classified as either intrinsic or extrinsic. The intrinsic membrane proteins are an integral part of the lens cell membrane, whereas extrinsic membrane proteins are associated only with the membrane surface. The principal intrinsic membrane protein is a 26,000-Da polypeptide, MIP-26, found in fibers but not epithelium.72–75 MIP-26 is abundant; it accounts for more than 60% of fiber membrane protein. Initially, MIP-26 was thought to be the unique protein component responsible for fiber-fiber coupling by gap junctions. However, subsequent studies have implicated connexin proteins in the formation of lens gap junctions. Connexin expression has been described previously. It appears MIP-26 may be a water channel.76 MIP-26 is, in fact, a member of the aquaporin family, AQP-O. Lens epithelium expresses a different, higher conductance, aquaporin, AQP-1.77 In the older lens there are increasing amounts of a 22,000-Da species of membrane protein (MIP-22).78 It has been suggested that MIP-22 is an age-dependent degradation product of MIP-26.78 Extrinsic membrane proteins have not been studied extensively in the lens. The presence of the glycoprotein fibronectin (MP 220–225) has been demonstrated. Found in many tissues, fibronectin is generally thought to be involved with the interaction between the cell surface and the extracellular matrix. A 43,000-Da (MP 43) extrinsic membrane protein is of some interest because its relationship with the lens cell membrane surface appears to be calcium dependent; this 43,000-Da polypeptide is thought to link aggregated crystallins to the cell membrane. The extrinsic membrane proteins of the lens are fully discussed by Alcala and Maisel.72 Many enzymes are associated with the cell membrane, including the transport enzymes Na, K-ATPase and Ca-ATPase, as well as alkaline phosphatase and adenyl cyclase.66,72 Enzymes can be either intrinsic or extrinsic membrane proteins. Glyceraldehyde-3-phosphate dehydrogenase is an extrinsic membrane protein (MP 37), whereas Na, K-ATPase (MP 90–107) is an intrinsic membrane protein. There are a number of membrane proteins that are associated with carbohydrates, predominantly galactose.79,80 Many of these membrane glycoproteins are intrinsic proteins associated with membrane receptors and surface recognition. CYTOSKELETAL PROTEINS This component of the lens water-insoluble protein fraction is soluble in urea. The microfilaments of the cytoskeleton are composed of actin found in two filamentous forms (β and γ) and in the globular form. The intermediate filaments are composed of a 57,000-Da molecular-weight polypeptide called vimentin. Beaded chain filaments, unique to the lens, consist of globular proteins attached to a backbone chain. Beaded filament composition may involve the expression of lens fiber-specific proteins CP49 and filensin. The microtubules are a complex structure containing two subunits of tubulin, each with a molecular weight of about 50,000 Da. The composition of the lens cytoskeleton has been reviewed by Quinlan and colleagues.81 AGING AND LENS PROTEINS The relative proportions of the various lens proteins change with aging.82 The amount of soluble protein fraction remains fairly constant with aging, but the insoluble protein fraction continues to increase. In the young lens the insoluble protein fraction might contribute only 1% of the total dry weight, whereas in a 70-year-old lens it can contribute more than 50%. This increase in the insoluble fraction is related primarily to a conversion of soluble protein into insoluble protein as a result of the formation of aggregates. The formation of large protein aggregates most probably contributes to a deterioration of lens transparency. With aging there is also an accumulation of low-molecular-weight polypeptides in the lens. These appear to be degradation products of crystallins as a result of proteolysis.83 ELECTROLYTES Because the lens is more or less a functional syncytium of tightly packed cells, the electrolyte composition of lens as a whole resembles that of a single cell. Relative to the surrounding environment of aqueous humor and vitreous, the lens has a high potassium content and a low sodium content.84 The potassium level is about 125 mmol/kg of lens water, and sodium is 14 to 26 mmol/kg of lens water. Lens chloride, which closely parallels lens sodium, is about 26 mmol/ kg of lens water.85 Cataractous lenses with any cortical involvement always manifest deranged electrolyte and water balance.86 This is not the case for nuclear cataracts. The divalent cation calcium is found at very low levels in the normal clear lens. Studies with calcium-sensitive fluorescent dyes suggest that most of the intracellular calcium is not freely diffusible; the free calcium concentration in lens cytoplasm is only about 100 nM.87 Calcium levels rise dramatically in most forms of cataract with cortical involvement.88,89 Normally, calcium is important in the maintenance of lens cell membrane permeability.90,91 Abnormally high concentrations of intracellular calcium are cytotoxic; they are likely to promote apoptosis (programmed cell death), to cause changes in protein aggregation, and almost certainly to contribute to the mechanism of cataractogenesis.92 Cytoplasmic pH in lens cells has been reported in the range 6.9 to 7.2, which is slightly more acidic than aqueous humor.93,94 The concentration of free amino acids in the mammalian lens is higher than that in the aqueous humor, vitreous, or plasma.95,96 There is good evidence that amino acids are transported into the lens by means of an energy-dependent carrier system. Little is known about the precise distribution of amino acids within the lens, but it is safe to assume that they are present at high levels in the epithelium and outer most fibers, the major site of protein synthesis. |
| LIPIDS | |||||||||||||||||||||||||
| The lipid content of the lens amounts to 1% to 2% of lens wet weight. Principally
associated with lens cell membranes, much of the lipid occurs
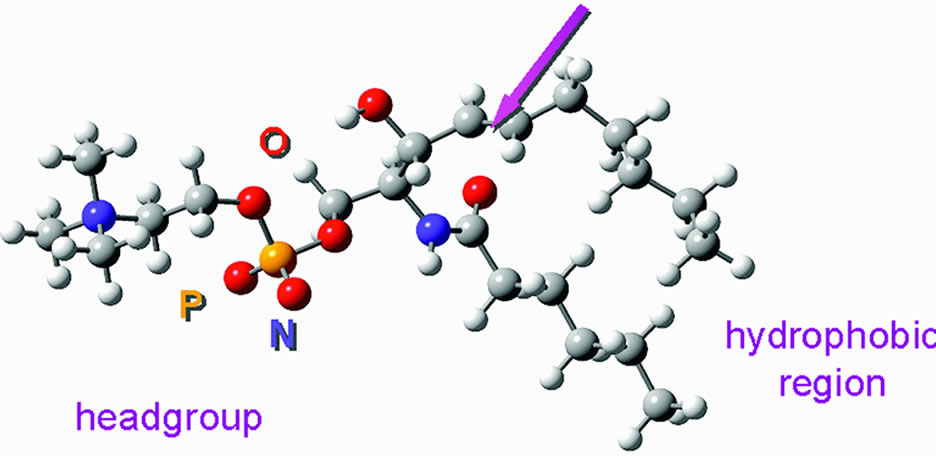
in a protein lipid complex. As reviewed by Zelenka,97 the lipids include cholesterol, phospholipids, and glycosphingolipids (Table 3). The major phospholipid in the human lens is dihydrosphingomyelin, which
is not abundant in any other tissue98 (Fig. 15). In the rabbit and bovine lens, phosphatidylethanolamine predominates.79 Cholesterol is abundant. It makes up about 50% of the total lens lipid; its
concentration is so great that the ratio of cholesterol to phospholipid
in a human lens is the highest known.99 This high cholesterol content together with the large amount of sphingolipid100 in the lens is consistent with the lens cell membranes being unusually
rigid; direct measurement of lens cell membrane lipid fluidity using
spectroscopic methods confirms this.101
Table 3. Phospholipid Composition of Lens Fiber Membranes
The values indicate the amount of each phospholipid as a percent of the total lens phospholipid. SPH, sphingomyelin; PC, phosphatidylcholine; PE, phosphatidylethanolamine; PS, phosphatidylserine; PI, phosphatidylinositol. The table is based on an original study by Zelenka (1984).
Substantial changes in lens lipid composition and distribution occur with aging. There is a doubling of lens cholesterol from about age 25 to 75 years and a concomitant increase in sphingomyelin; on the other hand, phosphatidylethanolamine and phosphatidylcholine levels decrease with aging. These changes, which reflect alteration in cell membrane structure, probably influence lens cell membrane function as the lens ages and may contribute to changes of ion permeability that occur with age.89 |
| ASCORBIC ACID |
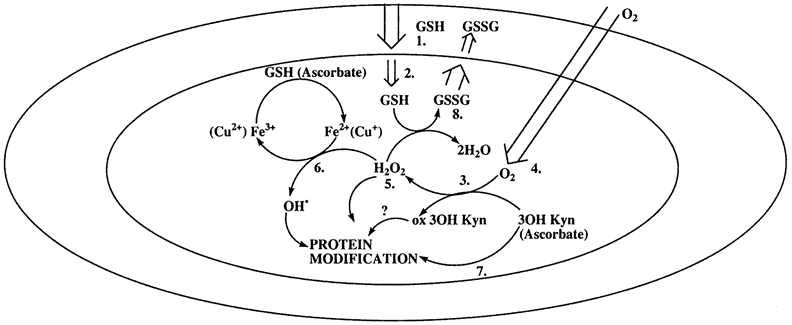
| In humans and many other species, ascorbic acid is found at high concentrations in the lens, sometimes higher than in aqueous humor.102 Ascorbic acid is not synthesized in the lens, but there is evidence that a specialized transport mechanism exists to transport it inward from the surrounding fluid.103,104 The role of ascorbic acid in the lens is the subject of some debate. It is likely to function as a scavenger of free oxygen radicals. Paradoxically, however, ascorbic acid might also be pro-oxidant, because together with light and the presence of a metal ion, ascorbic acid will generate hydrogen peroxide.105,106 Moreover, ascorbic acid might play a role in protein glycation and the development of chromophores that occur in older lenses. |
| GLUTATHIONE |
| Glutathione plays an important role in lenticular metabolism.107 There is a negligible quantity of glutathione in aqueous humor, but the levels of glutathione in the normal lens are very high. In the lens epithelium, glutathione has been reported at levels of 64 μmol/g wet weight; less is found in the lens cortex and nucleus. Mostly, lens glutathione is in a reduced state. Although glutathione is certainly synthesized by the lens, there is also evidence that the lens may have a glutathione transport mechanism.108 Glutathione levels fall significantly in cataract formation. The enzymes needed for the synthesis of glutathione may be present at a decreased level in the cataractous lens. |
| INOSITOL AND TAURINE |
| The levels of inositol and myo-inositol (the most abundant isomer of inositol) in the human lens are high. A sodium-dependent transporter enables the lens to accumulate myo-inositol from the aqueous humor.109 The lens appears to use myo-inositol as an osmolyte, accumulating more, or less, of the solute as a means of balancing cytoplasmic osmolarity with that of the surrounding fluid. Changes in expression of the myo-inositol transporter can cause lens opacification.110 Inositol is required as a precursor for membrane phosphoinositides and, thus, the maintenance of membrane integrity. Further discussion of the role of inositol is given by Dickerson and Lou.111 Taurine may also be an osmolyte,112 but it has antioxidant properties as well.113 Taurine is actively transported into the lens,114 in addition to being produced within the lens from methionine.115 |
| BIOCHEMISTRY | |
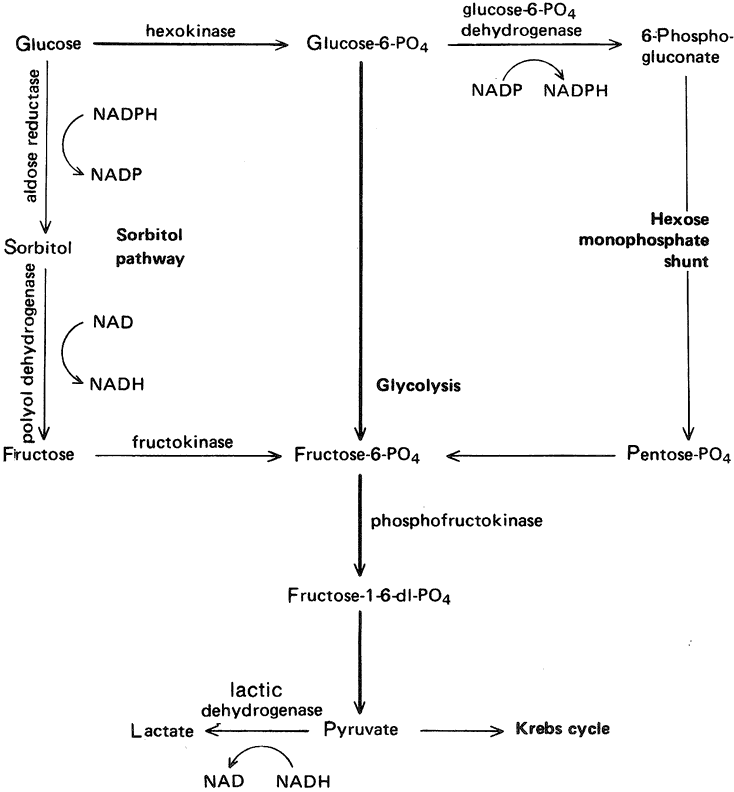
GLUCOSE METABOLISM The lens, like all cellular tissues, must generate a continual supply of ATP. The ATP is used to provide energy for processes such as active solute transport, synthesis of protein and lipid, and cell division. About 10% of the ATP generated is used for protein synthesis. The greatest need for energy in the lens appears to be within the epithelium. In the lens, as in most other tissues, a large fraction of the available ATP is used by Na, K-ATPase. Anaerobic glucose metabolism plays an important role in the lens (unlike most other tissues). Anaerobic glycolysis produces about two thirds of the ATP in the lens. Anaerobic glycolysis generates 2 moles of ATP for each mole of glucose. Although the enzymes necessary for glycolysis are found throughout the lens (see Cheng and Chylack116 for review), the first enzyme of the glycolytic pathway, hexokinase, which catalyzes the conversion of glucose to glucose-6-phosphate, is present at very low levels.117 The low level of hexokinase appears to be the rate-limiting factor for lens glucose metabolism and, therefore, restricts the generation of glucose-6-phosphate for both glycolysis and the hexose monophosphate shunt pathway118 (see later discussion). Anaerobic glycolysis, although not as efficient as the aerobic glucose metabolism process, obviates the problem of oxygen starvation in a tissue totally dependent on the aqueous humor, which has an unusually low oxygen tension. Indeed, lenses can survive under incubation conditions in the complete absence of oxygen as long as an adequate supply of glucose is available. However, when the lens is deprived of glucose, it rapidly uses up endogenous resources and begins to deteriorate, which results in loss of transparency. About 80% of the glucose entering the lens is converted to lactic acid by means of anaerobic glycolysis. Some of this lactic acid is metabolized further by the Krebs cycle, but most of it simply diffuses out into the aqueous humor to be eliminated from the eye. This explains why the aqueous leaving the eye has a much higher lactate concentration than newly formed aqueous humor.119 The aerobic metabolism of glucose is considerably more efficient than glycolysis, producing 38 moles of ATP from each mole of glucose. In the lens, however, ATP production by means of the Krebs cycle is mostly limited to the epithelium and perhaps some of the newly formed fiber cells at the lens equator. The epithelium possesses the necessary enzymes and probably has an adequate oxygen supply from the newly formed aqueous humor that flows across the anterior surface of the lens. It has been estimated that only about 3% of the total glucose is metabolized by means of the Krebs cycle, but because of the efficiency of the pathway this could generate up to 20% of the total ATP in the lens.120 Two other significant pathways of glucose metabolism operate in the lens: the hexose monophosphate shunt and the sorbitol pathway.121 Although these pathways do not generate a very large amount of ATP, they have received a lot of attention because of the relationship of nicotinamide adenine dinucleotide phosphate reduced form/nicotinamide adenine dinucleotide phosphate (NADPH/NADP) to glutathione metabolism and the link of the sorbitol pathway to sugar cataract. In the rabbit lens, about 14% of the glucose is metabolized by the hexose monophosphate shunt. This pathway uses glucose-6-phosphate as its initial substrate and generates pentoses, which are used in nucleic acid synthesis, and NADPH, which is an essential cofactor in many biochemical reactions (Fig. 16). NADPH is required for the maintenance of reduced glutathione by glutathione reductase and is also a necessary cofactor in the sorbitol pathway. Some of the pentoses are recycled to reenter the glycolytic pathway. Carbon dioxide produced by the hexose monophosphate shunt diffuses into the aqueous humor.
The sorbitol pathway of the lens converts glucose to sorbitol and then to fructose using the enzymes aldose reductase and polyol dehydrogenase, respectively (see Fig. 16). Under normal conditions, no more than about 5% of the glucose used by the lens is metabolized by the sorbitol pathway. Although it was first believed to be an ancillary ATP-generating mechanism, the sorbitol pathway has also been proposed as a means of protecting the lens from osmotic stress in hyperglycemia (see Cheng and Chylack116 for review). The important link between the sorbitol pathway and sugar cataract has caused researchers to focus on the possible role of aldose reductase in the cataractogenic mechanism.122 The availability of inhibitors for aldose reductase produced the encouraging finding that galactose cataracts in laboratory animals can be prevented by aldose reductase inhibition. However, the advances in the development of aldose reductase inhibitors for the treatment of human diabetic cataract have been complicated by the apparent differences between the human lens sorbitol pathway and sorbitol metabolism in animal lenses.116,122 PROTEIN METABOLISM Protein synthesis takes place mainly in the lens epithelium and superficial cortex.123 The process occurs slowly. Merriam and Kinsey124 estimated that glycine and serine turnover was no more than 5% per day. This makes sense when one considers that, once made, most lens proteins, at least in mature fibers cells, are retained for life. Specific crystallins are synthesized at different times during the development of the lens. The pattern of protein synthesis in the lens epithelium is different from that in lens fibers. α-Crystallin is expressed early in lens morphogenesis and characteristically appears in all lens cells. β- and γ-crystallins appear later in lens morphogenesis and are only synthesized by lens fibers.125 In the rat, epithelium and fibers express different isoforms of Na, K-ATPase.126 The expression of aquaporins and connexons is also different in fibers and epithelium.47,50 Using immunofluorescence techniques to study lens development, McAvoy and colleagues15 determined that β- and γ-crystallin synthesis can be detected when the epithelial cells differentiate and elongate to become lens fibers. Shestopalov and Bassnett54 demonstrated that fibers that retain their nucleus remain competent in their ability to synthesize proteins but this ability is probably lost once the fiber cell ages to the point when nucleus and other organelles are degraded. Because the proteins in the mature fiber cell remain in the lens for life, attention has been paid to the molecular modifications of the primary gene products that take place following their synthesis. There has been considerable study of the aggregation of lens proteins,127,128 because high-molecular-weight aggregates could cause light scattering and threaten the transparency of the lens. There have been a number of studies suggesting that, during the aging process, lens proteins may be susceptible to changes through oxidative mechanisms and other biochemical reactions such as phosphorylation and nonenzymatic glycation.129–131 The breakdown of obsolete or damaged proteins in the lens is catalyzed by peptidases and proteases. The lens contains a number of different endopeptidases, including neutral proteinases, and also the calcium-activated calpains.132 Proteins can also be broken down by a multicatalytic proteinase complex (proteasome).133 The turnover of proteins in the epithelium is consistent with the need for proteases. Their presence in lens fibers may be connected with fiber maturation, as well as with the degradation of proteins that are damaged during aging. GLUTATHIONE METABOLISM The finding of a large amount of glutathione in the lens, particularly in the epithelium and superficial cortex, together with the fact that the glutathione level is markedly reduced in cataractous lenses, has attracted a great deal of interest.107 One of the principal duties of glutathione is to maintain lens protein sulfhydryl groups (-SH) in the reduced state. Oxidation of lens protein sulfhydryl groups is unwanted because it leads to cross linkage of proteins, resulting in aggregation and loss of lens transparency.134 When the system works correctly, it is glutathione (GSH) that becomes oxidized, not the lens protein. Two molecules of oxidized glutathione link as G-S-S-G. The recycling of oxidized glutathione back to the reduced form is accomplished by a mechanism in which glutathione reductase couples the reduction of glutathione to the change of NADPH to NADP (see reviews in Spector129 and Augusteyn135). Glutathione may play a role in a number of other oxidation-reduction systems.135,136 For example, Giblin and coworkers137 have demonstrated a clear link between glutathione, glutathione peroxidase, the hexose monophosphate shunt, and hydrogen peroxide breakdown. In addition, glutathione metabolism is also linked to the removal of xenobiotics from the lens.108 |
| ELECTROLYTE BALANCE | |
| A balance between specialized solute transport processes and passive permeability
of lens membranes is necessary for the regulation of the electrolyte
and water content of the lens. In addition, specific transport
processes import nutrients. The exchange of substances between the
lens and aqueous humor is related to the properties of lens cell membranes
because solutes meet with little or no resistance from the capsule. A
large portion of the ATP in the lens is used to provide energy for
active transport mechanisms. The lens is able to concentrate such substances
as potassium, amino acids, glucose, taurine, and inositol, and
it actively extrudes solutes such as sodium and calcium. In addition, there
is a continual passive diffusional exchange of water, ions, and
waste products of metabolism, such as lactic acid and carbon dioxide. Maintenance
of proper cation balance in the lens is particularly important
because increased hydration of the cells results in cell swelling
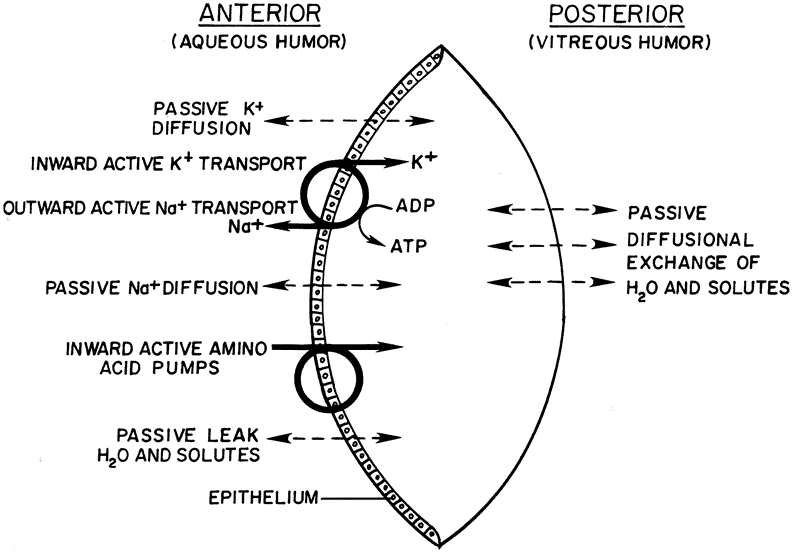
and this leads to a loss of lens transparency. In 1951, Harris and Gehrsitz presented some of the first evidence illustrating that the lens sodium and potassium levels are controlled by metabolically dependent transport processes.138 Refrigeration of rabbit lenses led to a fall in potassium content and an increase in sodium content; the cation redistribution could be reversed by warming the lens back to 37°C in a suitable culture medium. This recovery phenomenon can be reduced or abolished by metabolic inhibitors.84 The principal site of active cation transport in the lens is the epithelium, although sodium and calcium transport mechanisms are also present in the cortex.139–141 Curiously, the abundance of Na, K-ATPase protein in lens epithelium is similar to the abundance in fiber cells even though Na, K-ATPase activity in the fibers is much lower.142 Na, K-ATPase mRNA126 is detectable in the epithelium but not in the fibers. The dominant active ion transport mechanism is Na, K-ATPase, the sodium pump, which actively extrudes sodium and accumulates potassium. The ion gradients established by Na, K-ATPase provide the driving force for other processes such as sodium-calcium exchange, sodium-bicarbonate cotransport,143 and amino acid transport.144 The mechanistic details of the sodium pump are well documented.145 The combination of passive diffusion and anteriorly located energy-dependent transporters has been described as a “pump-leak” system146 (Fig. 17). Inhibition of Na, K-ATPase causes the lens to lose potassium and gain sodium. When this takes place, the osmotic equilibrium of the lens is disturbed and it swells as water enters the cytoplasm from the surrounding medium. Swelling and the consequent rupture of cells in the lens periphery obviously impair transparency. A number of investigators have explored whether the lens sodium pump has failed to operate in the human cortical cataract in which deranged lens electrolyte and water levels are a common finding.86 Although some investigators have detected a lower Na, K-ATPase activity in human cataractous lenses,147–149 others have reported no such difference.150,151 It has been suggested that lens epithelial cells are capable of synthesizing additional Na, K-ATPase protein to boost sodium pump activity in response to an increase of membrane permeability.152
Because high calcium levels are cytotoxic,92 and because the lens calcium level rises dramatically during the formation of cortical cataract, attention has been given to the mechanism by which a normal low concentration of calcium is maintained by the lens. It has been shown that the lens expresses calcium-stimulated ATPase, Ca-ATPase, both in the epithelium and cortical fibers.140,153 The plasma membrane-localized Ca-ATPase is capable of transporting calcium outward from the cytoplasm and plays a role in extruding calcium from the lens.139 Calcium is also exported via Na-Ca exchange, which couples the inward flow of sodium to the efflux of calcium. In addition, calcium is removed from the cytoplasm by a different Ca-ATPase, localized on the surface of ER.154 In the epithelium, calcium that has been shifted into the ER can be released as part of a signaling mechanism that is triggered when the lens is exposed to certain agonists.87,155 The specific permeability characteristics of lens membranes, which are clearly vital to the pump-leak mechanism of cytoplasmic ion regulation, hinge on the function of ion channels. Patch clamp studies have identified potassium conductance pathways near the lens surface. At least three different potassium channels contribute to potassium conductance; inward rectifiers, delayed rectifiers, and large-conductance calcium-activated channels.47 Chloride channels have also been detected, and it has been proposed that their function is essential to volume regulation of lens cells.156 Amino acids are accumulated by the lens, principally by the epithelium, and are therefore included in the pump-leak concept157 (see Fig. 17). There are a number of selective uptake mechanisms including one showing preference for alanine, another for leucine, and a third for glycine and other small amino acids.144 The sodium gradient between the lens cytoplasm and the surrounding fluid drives the amino acid accumulation mechanism; Marcantonio and Duncan158 have reported that the high sodium content of human cataractous lenses markedly reduces amino acid uptake. The uptake of glucose and other sugars has been discussed by a number of authors, including Kern.144 Unlike the active transfer of cations and amino acids, the transfer of glucose appears to occur across both the anterior and posterior surfaces of the lens; the mechanism appears to be insulin independent. Facilitative glucose transporters GLUT-1 and GLUT-3 are expressed in lens cells.159 Once a solute such as glucose enters the lens, a considerable time would be required for simple diffusion from the surface cells to the nucleus; in the human lens it has been calculated that 4 to 8 days would be necessary!47 However, it is thought that an inward movement of fluid occurs along the intercellular clefts at the anterior and posterior poles of the lens, and this would speed the penetration of glucose and other solutes into the lens cell mass. |
| CIRCULATION OF IONIC AND WATER FLUXES | |
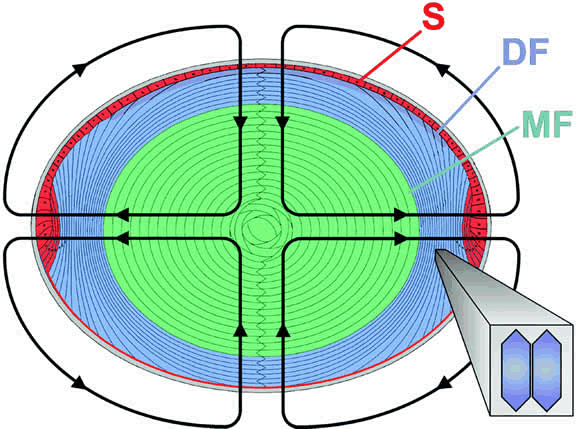
Mathias and his coworkers47 proposed the existence of circulating ionic currents that flow outward
at the lens equator and inward at the anterior and posterior pole (Fig. 18). Evidence for such currents was found initially by workers at Patterson's
laboratory160 who used a vibrating probe to detect electric current in the extracellular
fluid just outside the surface of the lens. The currents arise because
of the unequal spatial distribution of potassium channels, gap junctions, and
Na, K+-ATPase activity in the lens cell mass. Potassium channels are located
primarily in the membranes of cells at the surface, and the resting membrane
potential of surface lens fiber cells is considerably more negative
than that of fibers at deeper locations. Within the lens cell mass, the
radially oriented currents are likely to drive fluxes of ions, primarily
sodium, inward from the poles of the lens along intercellular
clefts. The loop circuit is completed by outward flow along a cell-to-cell
pathway via gap junctions that exit the lens at the equator. Such
an ionic circulation would drive a circulation of water, and this internal
circulatory system may enable the lens to overcome the difficulties
associated with the long diffusion time required for dissolved substances
to reach the center of the fiber mass. Solutes such as glucose
in the aqueous humor would tend to be shifted inward with the water
flow that enters the anterior pole and moves toward the center of the
lens along intercellular clefts.
|
| TRANSPARENCY |
| Lens transparency is made possible by a number of factors including the
regular arrangement of the lens fibers, the nonparticulate fiber cytoplasm, and
the uniform distribution and paracrystalline state of proteins
within the cells.161–163 Kuszak and colleagues164 have proposed that the arrangement of lens fibers depends strongly on
the ability of newly formed cells to elongate in a pattern that meshes
precisely with the underlying cells. The lamellar conformation of lens
proteins rather than helical structure may also contribute to transparency.165 In addition, it has been proposed that a short-range, liquid crystal-like
order of the crystallins is important for transparency of lens cytoplasm.166 It is not surprising, therefore, that lens proteins have the characteristic
of becoming opaque following phase separation at low temperatures167,168; this phenomenon contributes to cold cataract, the reversible opacification
of the lens when it is cooled. In the normal mammalian lens, body
temperature is higher than the critical temperature for protein phase
separation. The rainbow trout, which lives at cold temperatures, has
a lens that becomes reversibly opaque on warming.169 In addition to the state of lens crystallins, the tight packing of the lens cells and the regulation of ion and water balance also play significant roles in maintaining the transparency of the normal lens. Consequently, the development of protein aggregates, cell membrane degeneration, the appearance of vacuoles, and the distortion of lens structure can all produce light scatter and the clinical observation of cataract.162 It has been proposed that lens calcium and possibly pH levels may have an impact on the opacification process.92,170,171 Although the lens is transparent, the refractive index throughout the organ is not uniform. As a result of the more dense nature of the nucleus, it has a higher refractive index (1.41) than the cortex (1.38). The difference in refractive index between the nucleus and cortex results in the total refractive power of the lens being greater than if the refractive index were uniform throughout. Although from an anatomic standpoint, the cortex blends into the nucleus, distinct zones of optical discontinuity may be seen by slit lamp biomicroscopy. The zones are believed to correspond with various periods during the development and continual growth of the lens. |
| AGING OF THE LENS AND CATARACT | |
| With normal aging, lens weight and volume increase as the lens continues
to grow82,172 (see Figs. 6 and 7). As older cells are pushed toward the center of the lens, they lose their
nuclei and organelles. Protein synthesis and other metabolic processes
cease in the older lens cells. The lens develops a yellow color
with age. It can become dark brown as in brunescent cataract. The chemical
events leading to such color changes are discussed elsewhere.134,172,173 Also with aging, the lens becomes less elastic and this may hinder the
process of accommodation. The protein aggregation and other gradual molecular
changes that occur with aging almost certainly contribute to
a slow deterioration of optical quality. In many cases, however, the aging
process seems to be accelerated or metabolism is deranged to the
extent that there is a regional or widespread reduction in transparency
and vision is impaired. It is at this point that the clinical condition
of cataract exists. The incidence of cataract is high. In the Framingham Eye Study, which was restricted to a small sector of the population, it was found that detectable lens changes were observed in over 70% of all persons older than 65 years and that cataract was diagnosed in about 18% of that same age-group. In the age range 75 to 85 years the incidence of lens changes was above 90%, with senile cataract in about 46% of those studied.174 Within the United States it has been estimated that about 40,000 persons are legally blind from age-related cataract, representing 35% of existing visual impairment.175 More than 400,000 persons develop cataract each year. Cataract is a massive health problem worldwide; it is the greatest cause of global blindness. The World Health Organization estimated that 45 million persons throughout the world are blind, half of them as the result of cataract.176 Several studies have attempted to identify risk factors for cataract development.177–179 These include exposure to ultraviolet light, ionizing radiation, microwave radiation, toxic chemicals including prescription drugs, diabetes, elevated blood pressure, family history of high cataract prevalence, and a variety of inborn errors of metabolism. Several arguments have also been made to demonstrate a relationship between cataract and a variety of nutritional factors, including antioxidant intake. Recently the role of genetics in the development of cataract has come under scrutiny.180 Data from the Twin Eye Study suggest the relative influence of genetics and environmental factors on the development of human nuclear cataract is 48% and 14%, respectively.181 For cortical cataract, findings from the Twin Eye Study and the Beaver Dam Eye Study estimate that genetic factors could account for as much as 58% of cataract occurrence.182,183 The development of age-related cataract is obviously superimposed on the normal changes that take place in the aging lens. It is sometimes difficult to distinguish the alterations that normally occur with age from those lens changes that are precataractous. However, it is evident that the development of nuclear cataract and cortical cataract is not the same. Nuclear cataract is associated with coloration of the lens substance, but the lens electrolyte composition is unchanged134 (Fig. 19). In contrast, cortical cataract is almost always associated with deranged electrolyte and water balance.184 Lens sodium and calcium levels rise dramatically while there is a precipitous fall in lens potassium; this electrolyte imbalance causes an increase in cellular hydration and, thus, swelling. As might be expected, the development of cataract is accompanied by loss of the organized lens structure. Studies by Harding185 have demonstrated swollen and degenerate lens fibers, aberrant epithelial morphology and cell differentiation, and altered cell surfaces.
There has been extensive evaluation of changes in lens proteins associated with cataract.129,186,187 One certain fact is that with cataract development there is an abnormal increase in the fraction of water-insoluble protein in the lens and a decrease in total water-soluble proteins. The principal source of the increasing water-insoluble protein is the aggregation of soluble proteins to insoluble high-molecular-weight proteins. Aggregation of proteins also seems to be related to an increase in lenticular calcium. Some of the high-molecular-weight aggregates formed have been shown to be large enough to scatter light. However, it is unlikely that the formation of insoluble and high-molecular-weight proteins is alone sufficient to result in extensive lens opacification. It is known that some of the protein aggregates found in cataract are formed because of covalent disulfide bonds arising from the oxidation of thiol groups. In addition to altered membrane physiology and protein biochemistry, cataract development is accompanied by changes in lipid biochemistry,188 decreased enzyme activity,189 reduced glutathione levels,108 and diminished ATP production.190 Although the extensive alterations contribute to the demise of the cataractous lens, it is uncertain whether any single event initiates cataract development. There is an accumulation of evidence that suggests that oxidative damage might be one of the primary biochemical events leading to cataract development.191 In normal lenses there is only relatively minor oxidation, and this is principally associated with cell membranes. In cataract, however, there is considerable oxidation both at the level of the lens proteins and the cell membranes. If oxidation of lens constituents is indeed an initiating factor in cataract development, the potentially harmful species of oxygen responsible for the damage are superoxide, hydrogen peroxide, and the hydroxide radical.192 There is some evidence that the concentration of hydrogen peroxide is elevated in the aqueous humor of some patients with cataract.193 The source of hydrogen peroxide in the eye is uncertain, but ascorbic acid, present at normally high concentrations in the aqueous humor, can be a source of hydrogen peroxide in the presence of light and a metal ion. However, the lens normally has a number of mechanisms to protect it from oxidative damage, including catalase and glutathione peroxidase.108,135 Therefore, these mechanisms must be impaired or overcome if the lens is to become more susceptible to oxidative damage. Calcium might also play a significant role in eliciting molecular changes that would contribute to the loss of lens transparency. Elevation of calcium levels in cortical cataract was noted by Burge194 in 1909. A detailed discussion of calcium and cataract is given by Duncan and Jacob.88 Increased intracellular calcium levels can result in depressed glucose metabolism, inhibition of protein synthesis, induction of high-molecular-weight protein aggregates, and the activation of proteases, such as calpain92 and apoptosis. Calcium might directly inhibit the lens sodium pump,195,196 and intracellular calcium has been shown to modulate lens gap junction structure and coupling.197–199 Finally, elevation of cellular calcium may also be linked directly, from a physiochemical standpoint, to loss of transparency.199–201 |
| SUGAR CATARACT | |
| Although most cataracts appear to be age-related and are likely caused
by the combined effect of multiple factors, some instances of lens opacification
result from specific causes such as exposure to ionizing radiation,202 treatment with certain therapeutic drugs,203 and even as the consequence of being struck by lightning!204 Detailed information on cataract is available elsewhere, but it is informative
to consider aspects of sugar cataract. The two naturally occurring
forms of sugar cataract are diabetic cataract and galactosemic cataract. The
sorbitol pathway enzyme aldose reductase (see Fig. 16) plays a pivotal role in the process of cataract development. This enzyme
converts a sugar to its sugar alcohol, or polyol, which cannot readily
diffuse out of the intracellular compartment. Accumulation of the
sugar alcohol results in osmotic swelling, leading to opacification of
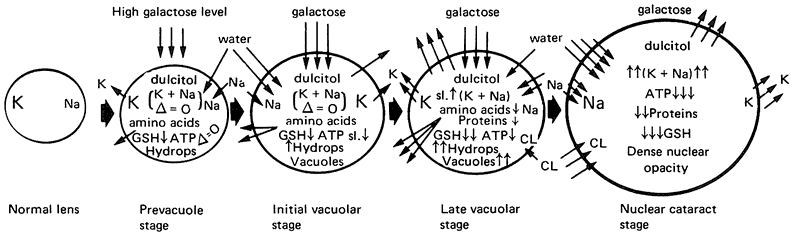
the lens.205,206 In diabetes mellitus the blood glucose level is elevated, resulting in an increase in aqueous humor glucose levels to nearly 90% of blood levels. As a result of the increased level of glucose in the aqueous humor, more glucose enters the lens by facilitated diffusion. In the normal lens virtually all the glucose entering is metabolized by way of hexokinase to enter the glycolytic pathway. However, at higher concentrations of glucose the enzyme aldose reductase is activated; this enzyme converts glucose to sorbitol, which accumulates within the fibers. Similarly, in galactosemia, a condition caused by deficiency of galactose-1-phosphate-uridyl transferase or galactokinase, galactose enters the lens and is metabolized through aldose reductase to dulcitol (galactitol). Dulcitol also accumulates within the lens fibers, causing an influx of water and swelling of the lens cells and eventually opacification (Fig. 20). Galactose cataract forms more rapidly than diabetic cataract because of the higher affinity of aldose reductase for galactose. The situation is worsened because the lens is unable to metabolize dulcitol. In contrast, sorbitol, the sugar alcohol of glucose, can be oxidized to fructose by the enzyme polyol dehydrogenase.
Sugar cataract can be induced in experimental animals by feeding galactose or by inducing diabetes. In studies with rats fed a diet of 50% galactose, the earliest visible change is the appearance of swollen peripheral lens fibers that eventually rupture, leading to water- and protein-filled clefts in the cortex.207 Later the lens epithelium proliferates into multiple layers. As the cortex becomes more liquefied, the lens temporarily attains a relatively clear appearance, but further changes result in a densely opaque nuclear zone, then total opacification. The accumulation of cellular water following polyol synthesis results in a number of further metabolic disturbances, including changes in membrane permeability, depletion of NADPH, reduction in glutathione levels, diminished ATP, and deranged electrolyte and balance.208 It has also been suggested that high levels of sugar may cause some direct modification of lens protein by a mechanism called nonenzymatic glycosylation.209 Glycosylation involves the attachment of glucose to amino acid residues in lens proteins, resulting in the formation of carboxyl groups that are able to cross link and, thus, cause protein aggregation. |
| CATARACT THERAPY |
| In North America, the approved treatment for cataract is surgical removal of the lens. In most cases, the optical quality of the eye is partially restored by implantation of a manmade intraocular lens. Presently there is no medical therapy for cataract available in North America but some anticataract drugs are marketed elsewhere in the world. Few of the drugs have been subjected to rigorous clinical trials to ascertain their efficacy. It is very difficult to design clinical studies on age-related cataract because lens opacification is hard to quantify and impossible to reverse and there is no clear-cut definition of precataractous lens changes. Anticataract preparations fall into various categories encompassing nutritional supplements (inorganic chemicals and vitamins), natural product extracts, aldose reductase inhibitors, antioxidants, and anti-inflammatory agents.210–213 The quest to develop an effective medical treatment for cataract is an important one. The development of a medical therapy that would delay the need for cataract surgery by just 10 years would have an enormous social and economic impact.214 The ability to develop this therapy clearly depends on the research effort to understand the risk factors and biochemical mechanisms responsible for the initiation and development of cataract. |
| ACKNOWLEDGMENTS |
| The manuscript was prepared with the expert assistance of Sara F. Byrd. The work was supported by NIH grant EY09532, an unrestricted grant from Research to Prevent Blindness, Inc., and the Kentucky Lions Eye Research Foundation. |