MONOCULAR VISION
Four separate characteristics of the light stimulus are identifiable: frequency and duration, intensity, the length of the light waves, and the angle of the light waves entering the eye.
Frequency and duration of the light stimulus are characteristics that evoke the on-off light sensation, providing the perception of a flickering or sustained light.
Intensity of the light stimulus is the characteristic that creates a light sensation ranging from weak to strong, providing perception of a dim or a bright light. The intensity of the stimulus is graded according to the volume of light waves that enters the eye per unit of surface area.
The length of the light waves is a characteristic within the visible spectrum that can be sorted out only by the cones of the retina and not by the rods. Regardless of the wavelength, the sensation of the rods is unvaried, and they perceive only light. The cones in photopic vision, however, respond with a multitude of varied sensations related to the wavelength of the light stimulus, each sensation providing varied color perception.
The angle of the light waves entering the eye is the characteristic of the stimulus that determines which retinal neuroepithelial photoreceptor is stimulated. Each stimulated photoreceptor has the innate ability to register the specific angle formed by the light wave and the corneal surface. This is the basis for spatially identifying the direction the light wave was traveling in reference to the eye at the time it entered. Photoreceptors in the fovea register a zero directional value, and the image projected here is perceived as centered in the visual field. A stimulated receptor nasal to the fovea localizes objects in space to the temporal side of center, a superior receptor localizes inferior to center, and so on. The number of degrees that each localization is away from center is equal to the number of degrees the stimulated neuroepithelial element is displaced from the fovea.
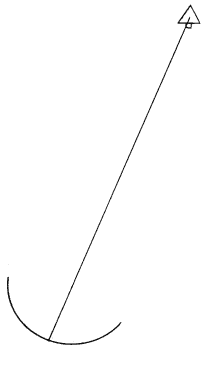
Since the refracting surfaces of the eye exert a constant effect on all penetrating light waves, the characteristic of the light wave angle—applied to an infinite number of light waves emanating from a multitude of light sources and comprising a patterned spatial stimulus—is the basis for the contoured image that projects onto the retina. Light waves emanating from objects in the visual field form miniaturized retinal images. The contour of each image is determined by the contrast produced between stimulated and unstimulated photoreceptors. The contrast sensation results in the perception of shape, size, and relative location of objects. Since the fovea possesses the highest resolving power for contrast sensation, projection of images onto this area results in maximal perception of contour, known as best visual acuity. The fixation point is the spatial location of the object of regard. The fixation line (visual axis) is a line drawn between the fixation point and the fovea (Fig. 1). The best visual acuity is obtained and maintained for the object of regard by a complex visuomotor reflex designed to steer the fixation line; this is known as the fixation reflex. It is an involuntary reflex mediated through the optomotor pathways, but it may be interrupted by disturbing innervations arriving at the extraocular muscles over the voluntary oculogyric pathways of the frontal lobe, the vestibular pathways, and the auditory and pain pathways.
|
The development of the fixation reflex is first manifest when the full-term infant is 4 to 5 weeks old. The eyes follow a light or a bright object for a few degrees, but once fixation is interrupted, reestablishment is slow. At 3 months of age, the infant maintains fixation in all fields of gaze and reestablishes fixation instantly after interruption. By 4 months of age, the infant begins to integrate fixation with grasping movements in an attempt to bring the target of fixation to the mouth. Oral identification of the object will continue for the next 4 months before being replaced by visual and tactile identification. By the time the child is 9 years old, the constant conditioning of the fixation reflex gives it an irreversible quality comparable to an unconditioned reflex; however, below this age, cessation of reinforcing this conditioning process results in its reversal, as manifest by amblyopia. If an infant is totally deprived of the opportunity to initiate stimulation of the fixation reflex before 3 months of age, the fixation reflex will never develop. Corrective treatment that eliminates the stimulus deprivation during approximately the first 3 months of life can facilitate fixation reflex development. At least this has been the clinical experience in treating patients with unilateral cataracts. Partial deprivation of stimuli, as occurs in strabismus and anisometropia, results in a less severe amblyopia and a relatively better prognosis for normal development of the fixation reflex with treatment. Treatment of strabismus amblyopia by 4 years of age is almost always successful, but the favorable prognosis decreases as the age at which treatment is initiated increases.
Absence of both maculas, bilateral congenital destructive disease of the maculas, or bilateral macular images below a critical threshold of intensity and clarity in infants results in a pendular nystagmus that first appears by 3 months of age. The nystagmus occurs when fixation is attempted, indicating a permanent loss of capability to steer and maintain the fixation line on the object of regard. This phenomenon suggests the degree of frustration associated with bilateral thwarted development of the fixation reflex. Deprivation nystagmus does not occur in patients with unilateral absence of the fixation reflex, even when their normal eye is occluded.
Best monocular visual acuity demands the combination of three essentials: a maturely developed fixation reflex that is normal in both its sensory and motor aspects, precise optical stimulation, and maximal attention directed to the object of regard. According to clinical observation, the fixation reflex appears to mature by the time the child is approximately 9 years old. Visual acuity remains in a state of flux until then, decreasing with disuse of the reflex and returning to normal with use. Sensory abnormalities of the reflex may reside in the retina or within the visual pathways. Abnormalities in the retina include abnormal anatomy, such as absent maculas associated with albinism, aniridia, and deficient cone physiology, as is found in achromatopsia. A lesion disrupting the macular fibers within the anterior visual pathways is detrimental to monocular acuity. Motor abnormalities of the reflex are exemplified by congenital motor nystagmus. Ptosis, opacities within the optical media, and abnormalities in refraction preclude precise stimulation of the fovea with intense and sharp images. Inattention to one of the foveal images (as occurs in strabismus), however, is the most common cause of normal visual acuity in an eye. Strabismus and absence of bifoveal vision (i.e., disregard of the image on one fovea) in the visually immature patient accounts for more amblyopia than all the above causes combined.
BINOCULAR VISION
Binocular vision is the unified perception that the visual cortex produces after integrating the separate sensations from the light stimulation of each eye. It is an acquired, conditioned reflex, having certain requisites for its development: (1) the bilateral retinal images must be similar in size, shape, and clarity; (2) these images projected onto bilateral retinal areas must have corresponding directional values; and (3) the above two requisites must be present by at least 2 years of age. Consequently, when these requisites are not satisfied, binocular vision does not develop in congenitally strabismic patients. The evidence is very convincing, however, that binocular vision will develop if the congenitally strabismic eyes are straightened during the critical period before 2 years of age.1–3 Grossly dissimilar images presented to each retina, as occurs in high degrees of anisometropia or in disturbances of the clear media of one eye, preclude the development of binocular vision, similar to congenital strabismus.
The first clinical evidence of binocular vision may be demonstrated by 6 months of age by adduction of the eye in response to an 8Δ base-out prism placed before it.
Since the cortex integrates the sensation produced by the light stimulus entering each eye into a single binocular perception, various characteristics of the light stimulus must be examined, and the cortical unification of their sensations should be assessed. The characteristics of the light stimulus that will be considered are the frequency and duration of the stimulus, the intensity of the stimulus, the wavelength of the light, and the angle the light entering the eye makes with the fixation axis.
FREQUENCY AND DURATION
Frequency and duration of the light stimulus in normal binocular viewing produces nothing different from monocular viewing. In laboratory viewing circumstances, however, when the eyes are alternately stimulated with flickering illumination in opposite cycles (i.e., one light comes on as the other goes off) and when cycled below the critical flicker-fusion rate, the result is binocular perception of a sustained light, but monocular perception of flickering light.
INTENSITY
Intensity of the light stimulus is also appreciated no differently in binocular versus monocular vision, except under rare circumstances. Two inadequate light stimuli directed separately into each eye are not additive in the sense that fusing the two can make one adequate stimulus. A dim light is not seen as brighter, nor is a bright light further intensified by seeing with two eyes rather than one. In fact, the binocular response is the same as the monocular response unless the light intensity entering each eye is different. In the laboratory, this situation can be produced by presenting a black, formless surface to one eye and a white, formless surface to the other. The fused perception is a lustrous metallic gray, ever changing by darkening and lightening in various portions as a result of retinal rivalry.
In the absence of binocular vision, this phenomenon does not occur, and the sustained impression of either a black or a white surface prevails. A similar phenomenon occurs in patients with a unilateral acquired cataract. When viewing a formless, illuminated background, such as the sky, the patient experiences a binocular dimness that clears on closing the cataractous eye and becomes dimmer on closing the normal eye; however, retinal rivalry persists when both eyes are open, manifest by the sky's increasing and decreasing in dimness in a patchy manner. This particular phenomenon is called interference.
WAVELENGTH
The length of the light wave under normal binocular viewing circumstances has little opportunity to yield a perception that differs from the monocular perception. The wavelength entering each eye, however, may be controlled by laboratory techniques; dissimilar lengths create some binocular perceptions that are totally different from monocular perception. Different colored lights presented to each eye are perceived either in color fusion, according to the scheme for mixture of colored lights rather than for mixture of pigments, or in color rivalry. Color fusion occurs only when very few combinations of colors of specific wavelengths are presented to the two eyes (Table 1). Otherwise, color rivalry of the two different formless fields of colored illumination occurs, with an ever-changing, patchy perception of the two. In the absence of binocular vision, either one field or the other persists as sustained, colored, and formless. Tints are also appreciated in patients with single binocular vision by presenting white light to one eye and a certain monochromatic light to the other. The diluted fused perception is a tint according to the scheme of mixing colored lights (see Table 1).
| Stimulus to One Eye | Stimulus to Other Eye | Fused Perception | |
| Complementary colors | Red | + Greenish blue | = White |
| Orange | + Cyan blue | = White | |
| Yellow | + Indigo blue | = Grayish white | |
| Greenish yellow | + Violet | = White | |
| Tints | Red | + White | = Pink |
| Orange | + White | = Yellow | |
| Yellow | + White | = Green | |
| Green | + White | = Yellow | |
| Violet | + White | = Salmon pink |
In sensory testing of a patient's binocular mechanism, color rivalry and color fusion (tints) form the basis of some tests. For example, a red lens before one eye and a green lens before the other causes the white viewed light to be perceived in color rivalry if the patient has single binocular vision; otherwise, either a red or a green sustained light is perceived. Another is the red glass test, in which a red glass is held before one eye while the patient views a white light. The patient with binocular vision sees a pink light (diluted from red), whereas the patient without fusion sees either a red or white light.
ANGLE
The angle of the light waves entering the eye has a significant response in binocular vision that is lacking in monocular vision. This is attributable to an object in space projecting onto retinal photoreceptors in each eye that have identical localizing values. A pair of such neuroepithelial elements is referred to as corresponding points. The foveas, as well as all other photoreceptors of the two eyes having identical directional values, are corresponding points. Images projected on these points are localized in space with the same values as though they were on only one of the corresponding points.
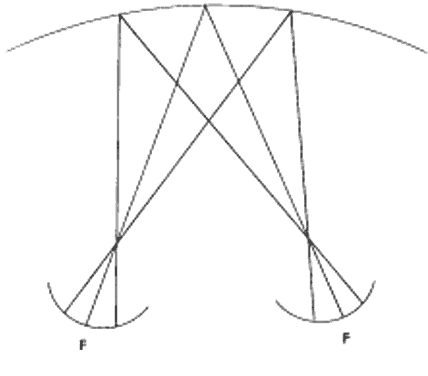
Lines drawn from corresponding points through the nodal points of the eyes intersect in space either at a distance remote from the eyes or at a distance close to the eyes, depending on whether the eyes are fixating a distant point or are converged to fixate a near point. An imaginary plane including all the points of intersection of these lines in space is a horopter (Fig. 2). The horopter is an infinitely thin toric plane in space with its center being the fixation point. The concavity of the distant horopter is almost nil, but it increases with increasing nearness of the fixation point and consequent convergence. An object located at any point on this plane projects images onto corresponding points on the retina. The angle formed by the intersecting fixating axes equals all the other angles formed by lines drawn from all other corresponding retinal points that intersect at the horopter.
|
Fusion
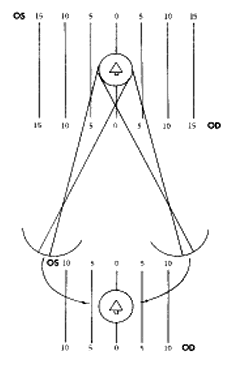
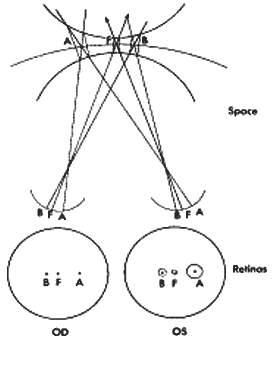
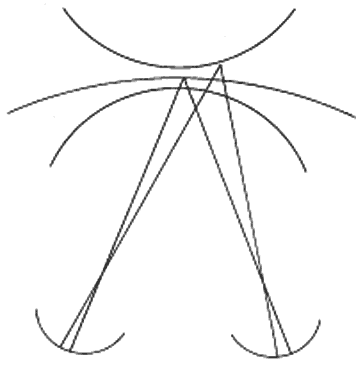
All objects projecting onto corresponding points have their two images fused into one perception; however, the neurophysiology involved in binocular vision does not impose such a rigid requirement for fusion. Objects projecting onto slightly disparate retinal points also may be fused, provided that the disparate images are within a critical limit. The quantity of disparity between foveal images that permits fusion is small, but the permissible disparity of images, which allows fusion, increases progressively for the more peripheral retinal points. Therefore, for each retinal point in an eye, there is a definable corresponding area in the retina of the opposite eye within which the same image must project if the two images are to be fused into one (Fig. 3). The more eccentric the retinal point from the fovea, the larger the corresponding area is in the other eye.
|
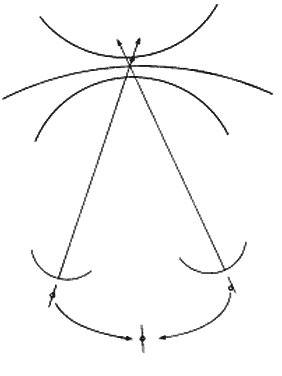
Lines drawn from the boundaries of the corresponding areas through the nodal point of the eye intersect in space, with each line extending from the corresponding point of the opposite eye. The line from the temporal boundary intersects the line of the corresponding point of the opposite eye proximal to the horopter (see Fig. 3). As in the construction of the horopter, if all points of the intersecting lines proximal to the horopter from retinal points and the temporal boundaries of the corresponding retinal areas of the opposite eye were connected by a plane, the proximal boundary of Panum's area would be defined. The distal border is defined by joining together all intersecting points distal to the horopter of lines from retinal points in an eye and the nasal boundaries of the corresponding retinal areas of the opposite eye. Since the peripheral retinal points have corresponding areas with larger dimensions than central retinal points, Panum's area is most narrow at its center and widens as it proceeds peripherally from the fixation area.
The retinal area in one eye that corresponds to the fovea in the other eye permits inexact alignment of the visual axes. While the object of regard is imaged on the fovea of one eye, it is slightly displaced from the fovea in the other, yet single binocular vision occurs rather than diplopic binocular vision. The inexact fixation of one eye (Fig. 4) in the presence of single binocular vision is called fixation disparity.4 It is physiologic, and the maximal quantity of deviation of the fixation axes may range between 10 and 14 minutes of arc. If there is a convergent deviation (esodeviation) of the eyes, although fusion keeps the deviation latent to the degree that single binocular vision is possible (esophoria), there is an esofixation disparity. Divergent deviation (exodeviation) is accompanied by exofixation disparity.
|
Simultaneous Perception
All objects distal or proximal to Panum's area project their images outside corresponding retinal areas and are not fused. An object outside Panum's area projects its images on the retinas with a degree of disparity exceeding the boundary of the retinal area of one eye that directionally corresponds to the retinal point receiving the image in the other eye. Although these disparate images are not fused, if they are perceived simultaneously, the object is simultaneously localized by the eyes to different places in space (diplopia). All objects not located within Panum's area are diplopic unless the observer is inattentive to these objects. The diplopia due to simultaneously perceiving objects distal and proximal to Panum's area is physiologic, since this phenomenon is a component of normal single binocular vision.
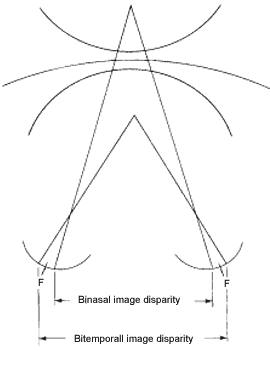
The physiologic diplopia is perceived as heteronymous for an object proximal to Panum's area, and homonymous for an object distal to Panum's area. If the object is in midline or laterally displaced short of a specific threshold from midline, the near object projects as bitemporal disparate images onto the retinas, and the distant object projects as binasal disparate images (Fig. 5). Laterally displaced objects beyond the specific threshold, however, project onto the temporal retina of one eye and onto the nasal retina of the other eye. This fact limits the usefulness of the terms “bitemporal disparity” and “binasal disparity” of the retinal images, yet these terms have been popularized in describing binocular parallax. A better and more realistic concept is to interrelate the nondisparate inter-retinal image distance from the object of regard with the disparate inter-retinal image distances from the objects distal and proximal to the object of regard. The more distal object always projects a less inter-retinal image distance than the object of regard, and the reverse is true for the nearer object (see Fig. 5).
|
Stereopsis
Within Panum's area, another single binocular vision phenomenon, stereopsis, occurs. Stereopsis is the perception of the third dimension (relative nearness and farness of object points within Panum's area) obtained from fusible but disparate retinal images. The same clues are used in stereopsis for determining the relative nearness and farness of object points within Panum's area as those used in physiologic diplopia to determine the same feature about object points proximal and distal to Panum's area. Thus, the stereopsis determination is made on the basis of differences in inter-retinal distances of the fusible images within Panum's area: the greater inter-retinal image distances are projected by nearer object points, and the lesser inter-retinal image distances are projected by further object points.
Fusion, stereopsis, and simultaneous perception are the three essential but distinctly different perceptual phenomena constituting single binocular vision. All three usually function simultaneously, although simultaneous perception is usually ignored unless a voluntary effort is made to recognize it. Only approximately 40% of congenitally esotropic patients—after their eyes have been straightened by surgery within the first 2 years of life and have developed simultaneous perception and fusion—ever develop stereopsis.5,6 Binocular vision is a cortical function with separate specialized cells for each of the following: simultaneous perception, fusion, and stereopsis. These are indeed considered to be three separate neurophysiologic phenomena; however, they all function only during the cortical processing of the images projected simultaneously onto each retina.
Terminology
Unfortunately, at the turn of the century, Worth7 used the term “fusion” for what is now called single binocular vision. He then introduced the terms “first-, second-, and third-degree fusion.” He described first-degree fusion as a simultaneous awareness of dissimilar targets presented in a haploscopic device (Fig. 6): for example, targets such as a fish to one eye and a bowl to the other (Fig. 7A). Second-degree fusion was the unifying of the similar portions of the two targets, each target having one minor dissimilarity (e.g., a flying insect with identical body but two sets of wings) in nonidentical positions. The body is fusible, but the wings are simultaneously perceived (Fig. 7B). Furthermore, the similar portions of each slide incited a motor response allowing fusional vergence amplitudes to be recorded; however, simultaneous awareness of dissimilar images on each retina has no capability of evoking a motor response. Third-degree fusion, as described by Worth, was the integrating of similar but disparate targets, obtaining a perception of stereopsis: for example, targets of a swing with the frame portion of the swing presented to each eye as nondisparate images, but the rope and the swing board suspended from the swing frame presenting either bitemporal or binasal retinal image disparity, making the rope and swing board appear swung toward or away from the swing frame when viewed in the haploscope.
|

|

Worth did not imply that all three degrees of fusion invariably occurred, but rather the following: (1) fusion may have been so rudimentary that only first-degree fusion was present; (2) second-degree fusion was better than only first-degree; and (3) if the patient possessed third-degree fusion, the fusion mechanism was excellent.
Gradually, Worth's terminology was altered, and the term “fusion” was replaced with the term single binocular vision. Worth's “first-degree fusion” became known as simultaneous perception; “second-degree fusion” was simply altered to fusion; and “third-degree fusion” was changed to stereopsis. However, many examiners who were indoctrinated with the haploscopic assessment of single binocular vision continue to think and articulate in terms of Worth's three levels of fusion, rather than the three perceptual phenomena that characterize single binocular vision.
The difficulties created by Worth's “three levels of fusion” concept start with the first level. The simple explanation of simultaneous perception as the ability simultaneously to perceive dissimilar slides presented to the viewer in a haploscope does not do justice to this complex neurophysiologic component of single binocular vision. Simultaneous perception of dissimilar images in a haploscope is the laboratory equivalent of only one of the two separate visual circumstances occurring in ordinary seeing that demonstrate simultaneous perception. The dissimilar targets in the haploscope simulate the circumstance that occurs in a nonstrabismic person who sees the visual environment at a distance by one eye and the obstruction precluding the same view by the other eye. Being aware of the distinctly different images simultaneously projecting onto the retinas is only one of the visual circumstances that reveals simultaneous perception. The other visual circumstance occurring during ordinary seeing that demonstrates simultaneous perception is attentiveness to an object outside Panum's area. This produces physiologic diplopia; that is, the similar images are not fused because they are too disparate. Worth's haploscopic dissimilar targets cannot produce this visual circumstance attendant with similar disparate images. Thus, his first-degree fusion test does not reveal the important neurophysiologic fact that physiologic diplopia is simultaneous perception of identical targets (objects) outside Panum's area.
Therefore, simultaneous perception occurs in response to two separate visual circumstances: (1) the appreciation of dissimilar images (Worth's first-degree fusion); and (2) the appreciation of similar images too disparate to fuse (physiologic diplopia). Simultaneous perception merely records the simultaneous spatial localizing values inherent for that moment within the retinal points of the two eyes when such points are presented with nonfusible visual stimuli, regardless of whether the visual stimuli are similar or dissimilar images.
The main difficulty with Worth's concept of fusion and stereopsis is that his terminology of second- and third-degree fusion does not impart the fact that fusion and stereopsis are separate perceptual responses to different visual stimuli arising only from within Panum's area. Although stereopsis is different from fusion, as suggested by the newer terminology, both occur simultaneously in normal single binocular vision. In fact, simultaneous perception, fusion, and stereopsis all occur simultaneously as three distinct phenomena constituting single binocular vision; they are not a system of grading the quality of single binocular vision, as Worth implied.
Since fusion and stereopsis both occur in response to visual stimuli arising from within Panum's area, there are some confusing ideas that they are one and the same neurophysiologic process; however, this is not the case. Each is a separate process requiring different stimuli and evoking different responses. Fusion of only one object point in an otherwise formless visual field (Fig. 8) can occur, but this is inadequate as a stimulus for stereopsis. Fusion of a multitude of object points arranged on a horopter can occur, but stereopsis of these object points is impossible. For stereopsis to occur, the retinas must be stimulated by at least two object points having a different proximal-distal relationship with each other in reference to the horopter (Fig. 9). Furthermore, the retinal image disparity produced by these object points must be horizontal, since stereopsis does not occur in response to vertical or torsional retinal image disparity. But fusion of all disparate retinal images that result from object points within Panum's area occurs, no matter whether the image disparity is horizontal, vertical, or torsional. Thus, all disparate retinal images producing stereopsis are also fused, but not all disparate images that are fusible produce stereopsis; also, nondisparate retinal images are fusible, but never will such images produce stereopsis.
|
|
One additional vast difference between fusion and stereopsis is the motor component. Fusion has a motor component; stereopsis has none. Fusional vergence is the motor component of fusion, designed to reduce horizontal, vertical, and torsional disparity of the retinal image to the degree that fusion can be attained or maintained. By fusing, stereopsis also is gained if the horizontal retinal image disparity is present. Stereopsis, however, has no identifiable isolated motor component designed solely to achieve the proper retinal image disparity that produces stereopsis. For example, the ideal circumstance for fusion is total absence of image disparity, and the fusional vergence aligns the eyes toward this goal, even though Panum's area makes it unnecessary to achieve it absolutely. Stereopsis, however, requires horizontal image disparity, but no motor component serves this requirement. Stereopsis appreciation becomes more and more difficult as the quantity of fusible retinal image disparity is reduced, and minimal disparity evoking this perception is 10 to 14 seconds of arc. Although in many everyday seeing circumstances stereopsis would be enhanced by adjusting the eye alignment and increasing the horizontal image disparity, it cannot occur because no motor reflex exists to produce it.
Still another difference between fusion and stereopsis is their respective effectiveness for distal object points. The horizontal retinal image disparity required for stereopsis is produced by the horizontal separation of right and left retinas. A separation of only approximately 6 cm soon becomes inadequate to yield sufficient horizontal retinal image disparity to produce stereopsis as the viewing distance between object points and the eyes is increased.
In 1838, Wheatstone wrote:
Although the accuracy of the stereoscopic sense is comparable with vernier acuity its range is comparatively limited. With the naked eye stereoscopic vision does not extend beyond 2000 feet. The radius of stereoscopic vision is about 7000 times the interpupillary baseline.
With the telestereoscope, invented by Helmholtz in 1857, the effective baseline was extended by the use of right-angled prisms. Prism binocular field glasses and stereoscopic range finders are constructed on this principle. In contrast to the limited range for stereopsis, fusion is equally effective for all ranges because the visual stimuli evoking it are not related to the interpupillary baseline.
Fusion and stereopsis serve different functions. Fusion localizes object points, in a two-dimensional plane for the observer, whereas stereopsis localizes in the third dimension of depth. Therefore, fusion and stereopsis differ not only in the visual stimuli required for their responses and in the nature of their responses, but also in their ultimate contribution to single binocular vision. They are quite different neurophysiologic entities, sharing only one component: the visual stimuli that produce their responses both arise from object points within Panum's area.